August 2024 TPJ Editor choice: Timing is everything: OsMYC2 controls diurnal flower-opening time in rice
Highlighted publication:
The OsMYC2–JA feedback loop regulates diurnal flower-opening time via cell wall loosening in rice
https://doi.org/10.1111/tpj.16910
Timing is everything: OsMYC2 controls diurnal flower-opening time in rice
Rice (Oryza sativa) is a major staple crop and the primary food source for over three billion people; thus, its yield is critical for global food security. Flowering is a key process affecting rice yield and is highly sensitive to temperature; in particular, high temperatures reduce pollen viability, severely impacting grain production. Consequently, flower opening in rice is adapted to its temperature environment. In the subspecies O. s. indica, commonly grown at lower latitudes, flowers open in the morning to avoid high temperatures in the middle of the day, whereas O. s. japonica, usually cultivated at higher latitudes, flowers around noon to avoid cold temperatures in the morning (Wang et al., 2019). Hybrids between indica and japonica often display strong heterosis effects, increasing yield by 30% compared to intra-subspecific indica hybrids. However, the non-overlapping flower opening times of the two subspecies restrict cross-pollination, making it difficult to implement cost-effective indica-japonica hybrid breeding programmes.
Flower opening in rice is initiated when the lodicules begin to swell, pushing the lemma and palea apart to expose the reproductive organs. The process requires rapid water absorption and expansion of the lodicule cells, controlled by the balance between the osmotic pressure and cell wall relaxation. The plant hormone jasmonate (JA) plays a crucial role in the process: treatment with methyl jasmonate as well as knock-out mutations of the JA signalling repressors JASMONATE ZIM DOMAIN 7 (OsJAZ7) and OsJAZ9 in the japonica cultivar Zhonghua 11 (ZH11) substantially advance flower opening time. How JA orchestrates diurnal flower opening downstream of these repressors, however, remains unknown.
Zhu and colleagues investigated the role of transcription factor OsMYC2, a homologue of Arabidopsis MYC2, which acts as a central hub of JA-controlled gene expression. They observed that OsMYC2 transcript levels in lodicules rose immediately before flower opening. Overexpression of OsMYC2 resulted in advancement of flower opening time from 11:00am to 9.30am and was linked to a shift in lodicule dynamics: lodicules size rapidly increased around 9.30am in OsMYC2 overexpressor (OsMYC2-OE) lines and had already started to shrink at 11:00am, when ZH11’s lodicules reached their maximum size and its flowers started to open (Figure 1). OsMYC2 overexpression did not strongly alter overall plant growth or other yield-related traits. OsMYC2-OEs and ZH11 did not differ in plant height, tiller number or seed setting rate, and only slightly increased grain numbers per spike and slightly reduced grain size were observed in OsMYC2-OEs.
The authors further investigated the molecular targets downstream of OsMYC2 by RNA-sequencing, identifying 2037 differentially expressed genes between OsMYC2-OE and ZH11. Genes with higher expression included many cell wall remodelling genes as well as the JA biosynthesis gene JASMONIC ACID CARBOXYL METHYLTRANSFERASE 1 (OsJMT1), and additional biosynthesis genes were found to be more highly expressed in lodicules when tested by qPCR. Nine of these genes harbour G- and E-boxes, canonical MYC2 binding sites, in their promoters, suggesting that these genes are directly regulated by OsMYC2. Using dual luciferase reporter assays and electrophoretic mobility shift assays (EMSAs), Zhu et al. showed that these genes are indeed bound and induced by OsMYC2.
What are the consequences of these changes in gene expression? Zhu et al. found substantially higher levels of both JA and its bioactive derivative JA-Isoleucine in OsMYC2-OE than in ZH11, suggesting a positive feedback regulation between OsMYC2 and JA biosynthesis. Their analysis also showed a lower cellulose and hemicellulose content in the lodicule cell wall in OsMYC2-OE than in ZH11, and this change was accompanied by thinner cell walls with decreased stiffness. These changes likely allow lodicules to more easily absorb water and expand, resulting in earlier flower opening. Taken together, the work by Zhu et al. uncovered a mechanistic link through which JA regulates cell wall relaxation to control diurnal flower-opening time.
Japonica and indica accessions harbour distinct haplotypes at the OsMYC2 locus, indicating a possible link between OsMYC2 alleles and the difference in flower opening times between these subspecies. In the future, the authors hope to leverage elite alleles of OsMYC2 and other diurnal flower opening time genes for cross breeding, improving cross-pollination and bringing down the cost of hybrid seed production.
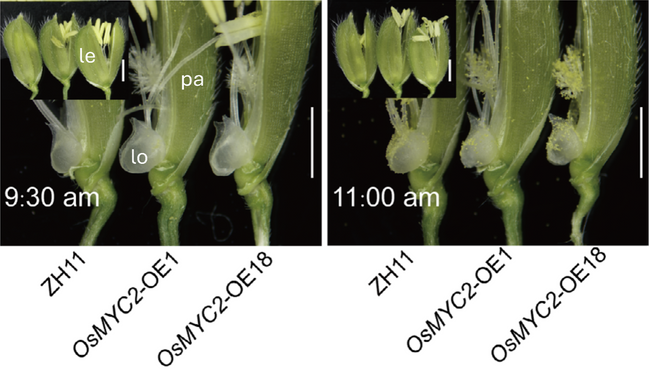
Figure 1. Effects of OsMYC2 overexpression on flower opening time
Changes in flower opening at 9:30am and 11:00am on the first day of flowering are depicted for the japonica cultivar ZH11 and two OsMYC2 overexpressor (OsMYC2-OE) lines grown under greenhouse conditions at South China Agricultural University. At 9:30am, enlarged lodicules (lo) in OsMYC2-OE trigger flower opening as they push lemma (le) and palea (pa) apart; at 11:00am, lodicules in OsMYC2-OE have started to shrink, while lodicules in ZH11 reach their maximum expansion and trigger flower opening. Scale bars represent 50 μm.
Figure modified from Zhu et al.